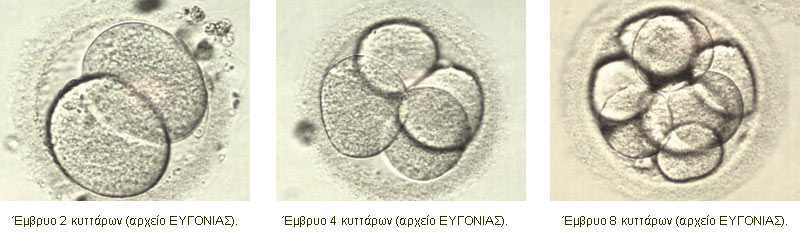
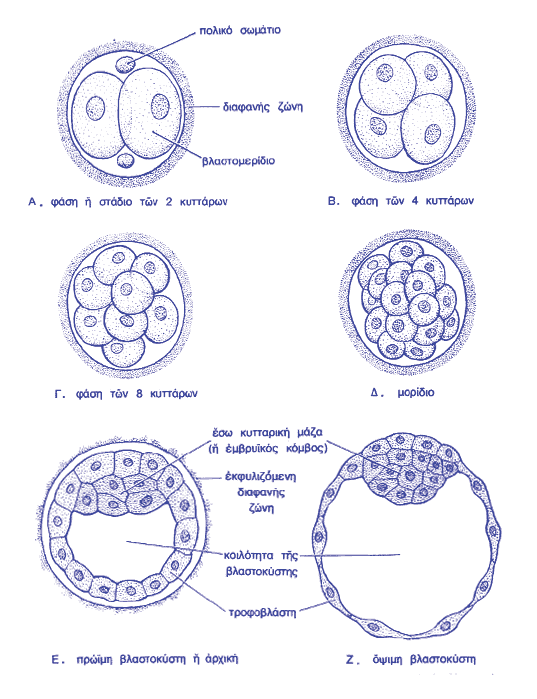
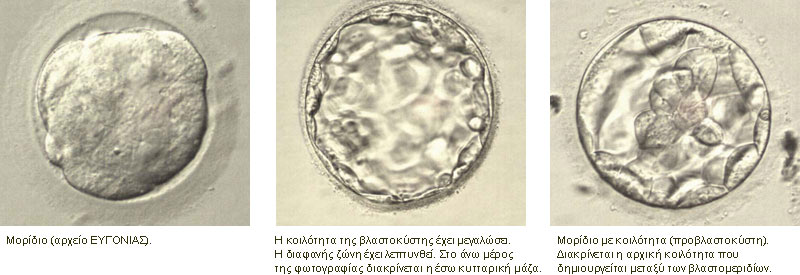
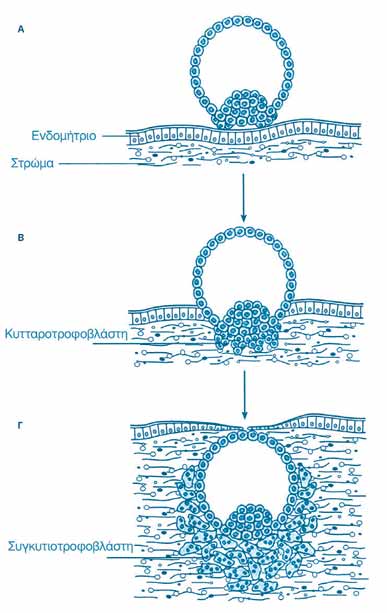
Σημείωση: το κείμενο που ακολουθεί βασίζεται σε αντίστοιχο κεφάλαιο από το βιβλίο του Χ. Ε. Καζλαρή «Το χρυσό μου παιδί», καθώς και σε διδακτικές σημειώσεις του ιδίου, με την ιδιότητα του Λέκτορα Εμβρυολογίας στο Τμήμα Ιατρικής του Πανεπιστημίου Θεσσαλίας. Οι τροποποιήσεις έχουν γίνει από τον συγγραφέα και το κείμενο αναδημοσιεύεται με την άδειά του.
Το κείμενο αυτό είναι απόσπασμα από το βιβλίο μας.
Μπορείτε να δείτε το σχετικό κεφάλαιο σε pdf εδώ:
ΣΥΝΟΠΤΙΚΗ ΙΣΤΟΡΙΑ ΕΜΒΡΥΟΛΟΓΙΑΣ & ΑΤΛΑΣ (3.4mb PDF) 
Η πρώτη ανθρώπινη αναφορά στη γονιμότητα θεωρείται ότι είναι η «Αφροδίτη του Willendorf», το αρχαιότερο γνωστό αγαλματίδιο εγκύου γυναίκας, που βρέθηκε στο ομώνυμο χωριό της Αυστρίας το 1908 (χρονολογείται στην περίοδο 24000-22000 π.Χ.). Πολύ αργότερα συντάσσονται τα πρώτα ιατρικά κείμενα, όπως ο πάπυρος Ebers (1550 π.Χ.), που θεωρείται το πρώτο γνωστό κείμενο με αναφορές στη γονιμότητα, αν και το κύριο αντικείμενό του είναι άλλο (περιέχει περίπου 800 συνταγές και αναφέρεται σε 700 φαρμακευτικές ουσίες).
Στην αρχαιοελληνική περίοδο, ο προσωκρατικός φιλόσοφος Ξενοφάνης ο Κολοφώνιος (περίπου 570-478 π.Χ.) εξετάζει απολιθώματα και εκφράζει απόψεις για την εξέλιξη της ζωής. Ο ιατρός Ιπποκράτης ο Κώος (460-370 π.Χ.) διατυπώνει πρώτος την αναγκαιότητα της επιστημονικής μεθοδολογίας και καταγράφει συστηματικές καθημερινές παρατηρήσεις της αναπτύξεως ωαρίων (αυγά όρνιθας), ενώ ο μεγάλος φιλόσοφος Αριστοτέλης (384-322 π.Χ.) συντάσσει πρώτος συγγράμματα φυσικής ιστορίας και συγκριτικής αναπτυξιακής ανατομικής. Για παράδειγμα, στα «Φυσικά» (350 π.Χ.) διατυπώνει κατ’ ουσίαν την επιγενετική θεωρία (το έμβρυο καθίσταται ολοένα και πιο πολύπλοκο κατά την ανάπτυξή του), θεωρία που έμελλε να αποδειχθεί ορθή μόλις τον 18ο αιώνα από τον Wolff (βλ. κατωτέρω). Ο Αριστοτέλης διατυπώνει επίσης θεμελιώδεις έννοιες της εμβρυολογίας, όπως τους τύπους εμβρυϊκής ανάπτυξης (ωοτοκία, όπου η γέννηση γίνεται από αυγά, όπως στα πτηνά, στα αμφίβια και στα ασπόνδυλα· ωοζωοζοκία, όπου τα αυγά εκκολάπτονται εντός του σώματος, όπως σε ορισμένα ερπετά και στους καρχαρίες· ζωοτοκία, όπου παρατηρείται γέννηση ζώντων νεογνών με πλακούντα, όπως στα θηλαστικά), τους τύπους αυλάκωσης των ζυγωτών (ολοβλαστική, όπως π.χ. στον βάτραχο και στα θηλαστικά, ή μεροβλαστική, όπως π.χ. στα πτηνά), τη λειτουργία του πλακούντα και του ομφαλίου λώρου, και πολλές άλλες. Ο Θεόφραστος από την Ερεσό της Λέσβου (371-287 π.Χ.), μαθητής του Αριστοτέλους, αρχίζει την πρώτη γνωστή συστηματική μελέτη της βοτανικής, ενώ ο Ηρόφιλος από τη Χαλκηδόνα της Βιθυνίας (περίπου 330-250 π.Χ.), ιατρός, μαθητής του Πραξαγόρα του Κώου, διενεργεί ανατομές, περιγράφει εσωτερικά όργανα του ανθρώπινου σώματος (οφθαλμό, εγκέφαλο, καρδιά, αρτηρίες-φλέβες, έντερο, γεννητικά όργανα) και δίδει τα ονόματά τους στον κερατοειδή και τον αμφιβληστροειδή χιτώνα και στο δωδεκαδάκτυλο. Ο Διοκλής ο Καρύστιος (240-180 π.Χ.), ιατρός, εισάγει τον όρο «ανατομία» και γράφει το πρώτο γνωστό σύγγραμμα ανατομικής.
Στους πρωτοχριστιανικούς χρόνους παρατηρείται κάμψη των επιστημών. Λίγα είναι γνωστά για την πειραματική επιστήμη της εποχής. Για παράδειγμα, αναφέρεται ότι ο Γαληνός (130-200 μ.Χ.) διενεργεί νεκροψίες χοίρων και συγγράφει βιβλίο ανατομικής του ανθρώπου. Ακολουθεί η μεγάλη σκοταδιστική περίοδος κατά την οποία χάνεται η αρχαία γνώση και οι αναφορές σε επιστημονικές παρατηρήσεις παραμένουν διάσπαρτες: για παράδειγμα, ο Mondino De Luzzi (1270-1326) διενεργεί νεκροψίες ανθρώπινων πτωμάτων και ο Guy de Chauliac (1300-1368) δημοσιεύει το πρώτο σύγγραμμα ανατομικής της «νέας» εποχής (“Chirurgia Magna”, 1363).
Στις αρχές της Αναγέννησης η επιστήμη αρχίζει να ανθίζει και πάλι. Ο Leonardo da Vinci (1452-1519), ο σημαντικός αυτός φιλόσοφος, αρχιτέκτων και βιολόγος, που εύλογα θεωρείται ως ο τελευταίος «πανεπιστήμων», διενεργεί τις πρώτες συστηματικές μετρήσεις της αναπτύξεως των ανθρώπινων εμβρύων in vivo, δημοσιεύει μάλιστα και ανατομικούς πίνακες με ρεαλιστικές απεικονίσεις από 30 νεκροψίες που πραγματοποιεί ο ίδιος (Εικ. 25Α). O Βέλγος ανατόμος André Vésale (Vesalius, 1514-1564) θεωρείται πατέρας της σύγχρονης Ανατομικής: διενεργεί συστηματικά νεκροψίες στο Πανεπιστήμιο της Padua και δημοσιεύει, το 1543, το σύγγραμμα “De humani corporis fabrica” (Περί της δομής του ανθρωπίνου σώματος), που από πολλούς θεωρείται το πιο σημαντικό σύγγραμμα της ιατρικής. Ο Ιταλός ανατόμος και εμβρυολόγος Hieronymus Fabricius ab Aquapendente (1553-1619) δημοσιεύει, το 1600, το σύγγραμμα “De Formato Foetu” (Περί του σχηματισμού του εμβρύου), που περιλαμβάνει τις πρώτες ρεαλιστικές απεικονίσεις εμβρύων. Ο Ιταλός ανατόμος Gabriele Fallopio (1523-1562) περιγράφει τον ωαγωγό, που φέρει σήμερα το όνομά του στην αγγλοσαξονική βιβλιογραφία (“fallopian tube”).
Η επιστήμη αρχίζει να αποκτά ολοένα και μεγαλύτερη αποδοχή, αλλά παράλληλα αρχίζουν και οι διαμάχες. Χαρακτηριστικό παράδειγμα είναι η ανακάλυψη της κυκλοφορίας του αίματος από τον μεγάλο Βρετανό ιατρό William Harvey (1578-1657): το 1628 δημοσιεύεται το σύγγραμμά του “Excercitatio Anatomica de Motu Cordis et Sanguinis in Animali” (Περί της ανατομικής της κυκλοφορίας του αίματος στα έμβια) και το 1651 το σύγγραμμα “Exertationes de Generatione Animalium” (Περί της δημιουργίας των εμβίων), στο οποίο διαπιστώνεται η αρχή ότι όλα τα ζώα προέρχονται από ωάρια (“ex ovo omnia” ή “omne vivum ex ovo”). Οι θεωρίες αυτές έρχονται σε αντίθεση με τον Αριστοτέλη, που πίστευε ότι τα υγρά της εμμήνου ρύσεως περιέχουν την ύλη του εμβρύου, στην οποία δίνει μορφή και ψυχή το σπέρμα. Στο δεύτερο βιβλίο του, ο Harvey περιγράφει πρώτος το βλαστόδερμα, την περιοχή του ωαρίου της όρνιθας από την οποία αναπτύσσεται το έμβρυο. Την ίδια περίπου εποχή, ο Ολλανδός φυσιολόγος, ανατόμος και εμβρυολόγος Reinier de Graaf (1641-1673) περιγράφει το ωοθυλάκιο στο κουνέλι (1672), μαζί με τον Niels Steensen (1638-1686). Ο σημαντικός αυτός Δανό ανατόμος ανακαλύπτει επίσης τον πόρο της παρωτίδος, γίνεται μετέπειτα γνωστός ως γεωλόγος (πατέρας της στρωματογραφίας) με το εκλατινισμένο όνομα Nicolaus Steno και τελικώς αποσύρεται από την επιστήμη, αφιερώνεται στην εκκλησία και γίνεται επίσκοπος (αγιοποιήθηκε μάλιστα από τον Πάπα Ιωάννη-Παύλο Β’ το 1988).
Ο 17ος αιώνας είναι ο αιώνας σημαντικών επιστημονικών επιτευγμάτων. Ένας Ολλανδός έμπορος, ο Zacharias Jansen (1632-1723) μόλις έχει εφεύρει το μικροσκόπιο (1595), συσκευή με την οποία ο Ολλανδός φυσιοδίφης Jan Swammerdam (1637-1680) παρατηρεί τα ερυθρά αιμοσφαίρια, το 1658. Ο Βρετανός φυσιοδίφης Robert Hooke (1635-1703) περιγράφει τα κύτταρα (σε φελλό, το 1663) και εκδίδει, το 1665 τη “Micrographia” (Μικρογραφία), το πρώτο σύγγραμμα ιστολογίας. Ο Ολλανδός έμπορος Antonie Van Leeuwenhoek (1632-1723) τελειοποιεί το μικροσκόπιο του Jansen, αυξάνοντος τη μεγεθυντική του ικανότητα (×300) και εισάγοντας τον κοχλία εστιάσεως. Ο Van Leeuwenhoek περιγράφει πρώτος τα μικρόβια (1674) και μαζί με τον Hamm περιγράφει τα σπερματοζωάρια (1677-1678). Ο de Graaf, που είναι φίλος του, τον εισάγει στη Βασιλική Εταιρεία του Λονδίνου. Ο Van Leeuwenhoek θεωρεί ότι στο σπερματοζωάριο υπάρχουν μυς και αρθρώσεις που εξασφαλίζουν την κινητικότητα. Σε ένα χαρακτηριστικό και εξαίρετο δείγμα επιστημονικής αμφισβήτησης, ο Van Leeuwenhoek γράφει μια φράση κλειδί, το 1717: “I know very well that there are Universities who do not believe that living creatures are in the male semen; but I do not mind about this, as I know I have the truth.” (Γνωρίζω καλά ότι, σε ορισμένα Πανεπιστήμια, η παρουσία εμβίων όντων στο σπέρμα των αρρένων δεν είναι παραδεκτή. Αυτό όμως δεν με απασχολεί, διότι γνωρίζω ότι κατέχω την αλήθεια.) Οι σύγχρονοί του «σπερματιστές» (με πρώτο τον Nicolas Hartsoeker, το 1694) βλέπουν μέσα στα σπερματοζωάρια μικροσκοπικούς ανθρώπους (Εικ. 25Β), τα «ανθρωπάρια» (“homunculi”: η επικρατούσα άποψη ότι ο Van Leeuwenhoek τα περιέγραψε είναι λανθασμένη). Ο Ιταλός ιατρός και βιολόγος Marcello Malpighi (1628-1694), μαθητής του Γαλιλαίου, θεωρείται ως ο πατέρας της Ιστολογίας. Το 1672 δημοσιεύει την πρώτη μικροσκοπική μελέτη της εμβρυϊκής ανάπτυξης της όρνιθας (στην οποία περιγράφονται η νευρική αύλακα, οι σωμίτες, η κυκλοφορία του αίματος, κ.λπ.) και το 1674 μια συγκριτική περιγραφή των εμβρύων των φυτών και των πτηνών. Αρχίζει μεγάλη θεωρητική διαμάχη μεταξύ των οπαδών της επιγένεσης και των οπαδών του προσχηματισμού. Το 1678, ο Malpighi δημοσιεύει περιγραφή των αδένων και των σπλάγχνων. Την ίδια εποχή, ο Βρετανός μαιευτήρας William Smellie (1697-1763) είναι ο πρώτος που διδάσκει συστηματικά τη Μαιευτική, αποσυνδέοντάς την από τη χειρουργική, επινοεί μαιευτικά εργαλεία και δραστηριοποιείται για την αναγνώριση της ιδιότητας του μαιευτή. Την περίοδο 1752-1754, ο Smellie δημοσιεύει συγγράμματα μαιευτικής με μοναδικής ακρίβειας απεικονίσεις εμβρύων σε πίνακες (“Smellie tables”).
Ο Ιταλός ιερωμένος και βιολόγος Lazzaro Spallanzani (1729-1799) αποδεικνύει, το 1775, ότι σπερματοζωάρια και ωάρια είναι εξ ίσου απαραίτητα για την αναπαραγωγή και λύει έτσι μερικώς τη διαμάχη μεταξύ «ωαριστών» (επιγενετική θεωρία) και «σπερματιστών» (θεωρία του προσχηματισμού). Το 1776, ο Spallanzani, σε ένα καταλυτικό πείραμα, θεμελιώνει την κρυοβιολογία: καταψύχει σπέρμα χρησιμοποιώντας χιόνι και παρατηρεί τη μείωση της κινητικότητος των σπερματοζωαρίων στο ψύχος. Το 1780 πραγματοποιεί και την πρώτη τεχνητή γονιμοποίηση (σπερματέγχυση) στον σκύλο, με τοποθέτηση σπέρματος στον κόλπο του θηλυκού. Λίγα χρόνια αργότερα, η νέα αυτή μέθοδος εφαρμόζεται για πρώτη φορά στον άνθρωπο από τον Βρετανό χειρουργό και ανατόμο John Hunter (1728-1793): το 1791 ο Hunter πραγματοποιεί, στο Λονδίνο, την πρώτη επιτυχή τεχνητή γονιμοποίηση (ομόλογο σπερματέγχυση) σε ασθενή.
Ο Γερμανός ανατόμος, βιολόγος και εμβρυολόγος Kaspar Friedrich Wolff (1733-1794) είναι κατά πολλούς η πιο σημαντική μορφή της επιστήμης της εμβρυολογίας μετά τον Αριστοτέλη. Ο Wolff πρακτικά αποδεικνύει την ορθότητα της επιγενετικής θεωρίας: περιγράφει συστηματικά τα στάδια της εμβρυϊκής αναπτύξεως στα πτηνά και διατυπώνει (1759) την ιδέα της θεωρίας των τριών δερμάτων για τα έμβρυα των σπονδυλωτών στην εργασία του “Theoria generationis” (Θεωρία της δημιουργίας), για την οποία δέχεται αυστηρότατη κριτική από το πανεπιστημιακό κατεστημένο της εποχής. Πράγματι, η θεωρία του προσχηματισμού, εάν ίσχυε, θα έπρεπε να αποκλείει την εξέλιξη των όντων, αφού θα υπήρχε «εγκιβωτισμός» (“emboîtement”) της κάθε γενεάς στην προηγούμενη. Όσοι επέμεναν να «βλέπουν» ανθρωπάρια μέσα στα σπερματοζωάρια, το έκαναν επειδή δεν είχε ακόμη διατυπωθεί η κυτταρική θεωρία, σύμφωνα με την οποία υπάρχει ένα ανυπέρβλητο όριο στον εγκιβωτισμό, το φυσικό μέγεθος των κυττάρων! Ο Wolff απέδειξε αντιθέτως ότι τα αγγεία αναπτύσσονται εκ νέου σε κάθε έμβρυο και ότι το έντερο σχηματίζεται από την πτύχωση ενός αρχικώς επίπεδου ιστού (1767): κατά συνέπεια, η επιγενετική θεωρία ισχύει, το έμβρυο όντως καθίσταται ολοένα και πιο πολύπλοκο καθώς αναπτύσσεται.
Παράλληλα, οι επιστήμονες στράφηκαν και στη μελέτη της εξέλιξης. Ο Γάλλος βιολόγος Jean-Baptiste Pierre Antoine de Monet, Chevalier de Lamarck (1744-1829) υπήρξε ο πρώτος που πρότεινε μια θεωρία της εξέλιξης, τη θεωρία του «μετασχηματισμού» (η εξέλιξη ενός είδους επέρχεται ως «μια νέα ανάγκη που εξακολουθεί να εμφανίζεται»). Σύμφωνα με τη θεωρία του Lamarck (1809), τα επίκτητα χαρακτηριστικά, τα οποία ο οργανισμός αποκτά κατά τη διάρκεια της ζωής του, κληροδοτούνται στους απογόνους του. Η θεωρία αυτή, ως γνωστόν, δεν ισχύει, ενώ αντιθέτως ισχύει η θεωρία της φυσικής επιλογής των Darwin και Wallace, που διατυπώθηκε όμως πολύ αργότερα (βλ. παρακάτω).
Η μελέτη της εμβρυολογίας αρχίζει να γίνεται συστηματική. Ο Γερμανός ιατρός, φυσιολόγος και νευροανατόμος Karl Friedrich Burdach (1776-1847) εισάγει, το 1800, τον όρο “βιολογία”, τον οποίο ο Lamarck αποδέχεται αμέσως. Ο Ελβετός βιολόγος Jean-Louis Prevost (1780-1850) και ο Γάλλος χημικός Jean-Baptiste Dumas (1800-1884) περιγράφουν (1824) την αυλάκωση σε έμβρυα βατράχου Rana esculenta και προτείνουν ότι τα σπερματοζωάρια δεν είναι παράσιτα, αλλά χρήσιμοι παράγοντες για τη γονιμοποίηση. Η ανακάλυψή τους περνά σχεδόν απαρατήρητη. Ο Γερμανός ανατόμος Martin Heinrich Rathke (1793-1860) μελετά την ανάπτυξη πολλών ειδών (βατράχου, σαλαμάνδρας, ιχθύων, πτηνών και θηλαστικών) και παρατηρεί ότι όλα τα σπονδυλωτά εμφανίζουν παρόμοια σχέδια ανάπτυξης. Σημαντική ανακάλυψη του Rathke είναι τα βραγχιακά ή φαρυγγικά τόξα (εξελίσσονται σε βράγχια στους ιχθείς, σε γνάθους και ώτα στα θηλαστικά). Ο Λεττονός ανατόμος Christian Heinrich Pander (1794-1865) μελετά την εμβρυϊκή ανάπτυξη της όρνιθας για μόλις 15 μήνες και ανακαλύπτει τα τρία βλαστικά δέρματα (1817), καθώς επίσης και ότι τα τρία δέρματα δεν δημιουργούν τους ιστούς και τα όργανα ανεξάρτητα, αλλά αντιθέτως, ότι υπάρχουν αλληλεπιδράσεις μεταξύ τους (αυτό το φαινόμενο θα ονομασθεί αργότερα “επαγωγή”).
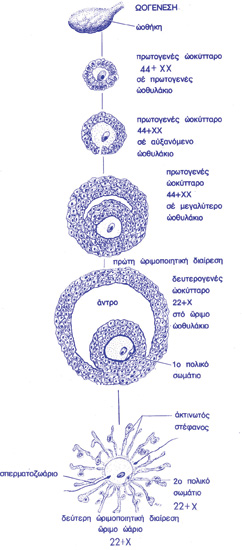
Πατέρας της σύγχρονης εμβρυολογίας θεωρείται ο Εσθονός εμβρυολόγος Karl Ernst von Baer (1792-1876), μαθητής του Burdach και φίλος των Rathke και Pander. Ο von Baer ανακαλύπτει τη νωτιαία χορδή, η οποία αποτελείται από μεσόδερμα και επάγει τη μετατροπή του εξωδέρματος σε νευρικό ιστό. Επί πλέον, αποδεικνύει την ύπαρξη του ωαρίου στα θηλαστικά, ονομάζει τα κινούμενα κύτταρα του σπέρματος σπερματοζωάρια», διατυπώνει τέσσερις «Αρχές της Εμβρυολογίας» και εισάγει τον κλάδο της συγκριτικής εμβρυολογίας (1837, “Uber die Entwickiunssseschichte der Thiere, Beobachtuns und Reflexion”). Οι τέσσερις θεμελιώσεις αρχές της εμβρυολογίας έχουν ως εξής:
Κατά την εμβρυϊκή ανάπτυξη, τα γενικά χαρακτηριστικά μιας μεγάλης ομάδας ζώων εμφανίζονται νωρίτερα από τα ειδικά χαρακτηριστικά μιας μικρότερης ομάδας ζώων (για παράδειγμα, τα έμβρυα όλων των σπονδυλωτών διαθέτουν φαρυγγικά τόξα, νωτιαία χορδή, σπονδυλική στήλη και αρχέγονο νεφρό).
- Τα λιγότερο γενικά χαρακτηριστικά απορρέουν από τα πιο γενικά, έως ότου εμφανισθούν τα πιο ειδικά (για παράδειγμα, η ανάπτυξη των άκρων είναι παρόμοια στην αρχή, τα άκρα εξειδικεύονται αργότερα).
- Το έμβρυο ενός είδους δεν περνά από τα στάδια ανάπτυξης των κατωτέρων (δηλαδή απλουστέρων) ειδών, αλλά αποκλίνει προοδευτικά από αυτά.
- Επομένως, το έμβρυο ενός ανωτέρου ζώου δεν μοιάζει ποτέ με ένα κατώτερο ζώο, αλλά με μια πρώιμη μορφή του εμβρύου του: για παράδειγμα, το ανθρώπινο έμβρυο δεν περνά ποτέ από κάποιο στάδιο ώριμου ψαριού, ενώ τα πρώιμα έμβρυα τους είναι παρόμοια.
Η επόμενη μεγάλη καμπή είναι η διατύπωση της κυτταρικής θεωρίας, η οποία αποδίδεται στον Γερμανό ζωολόγο Theodor Schwann (1810-1882), στον Γάλλο βοτανολόγο René Joachim Henri Dutrochet (1776-1847) και στον Γερμανό βοτανολόγο Matthias Jacob Schleiden (1804-1881), ο οποίος μεταξύ άλλων ανακάλυψε την όσμωση. Σύμφωνα με την κυτταρική θεωρία (1839) όλα τα φυτά και τα ζώα αποτελούνται από κύτταρα. Ο Schwann θεωρεί τους πυρήνες ως οριστικό κριτήριο περιγραφής των ζωικών κυττάρων (τους πυρήνες είχε διακρίνει πρώτος ο Βρετανός βοτανολόγος Robert Brown, το 1833). Την ίδια εποχή, οι δύο Βρετανοί βιολόγοι Charles Darwin (Δαρβίνος, 1809-1882) και Alfred Russel Wallace (1809-1882) διατυπώνουν, το 1858, τη «Θεωρία της Φυσικής Επιλογής των Βιολογικών Ειδών». Τα κυριότερα στοιχεία της θεωρίας αυτής βρίσκονται στο πολύκροτο σύγγραμμα “On the Origin of Species” (Περί της καταγωγής των ειδών). Μια από τις πιο σημαντικές ιδέες της δαρβινικής θεωρίας σχετικά με την εμβρυολογία είναι ότι η ομοιότητα των εμβρυϊκών δομών αντικατοπτρίζει την ομοιότητα των προγόνων (“community of embryonic structure reveals community of descent”). Τα παραλειπόμενα της ιστορίας είναι εν πολλοίς γνωστά: το 1858, ο Wallace στέλνει στον Darwin μια μελέτη του, στην οποία περιγράφει τη φυσική επιλογή, χωρίς να γνωρίζει ότι εκείνος ήδη μελετούσε μια παρόμοια θεωρία από εικοσαετίας, απομονωμένος στην έπαυλή του μετά τα ταξίδια του στις νήσους Galapagos, όπου είχε συλλέξει όλες τις απαραίτητες πληροφορίες και χιλιάδες δείγματα. Ο Darwin ολοκληρώνει εσπευσμένα τη δική του μελέτη και παρουσιάζει μια κοινή ανακοίνωση της νέας θεωρίας στη Λινναία Εταιρεία (Linnaean Society) την 1η Ιουλίου 1858.
Στον πειραματικό τομέα, ο σημαντικότατος Πρώσσος ιατρός και ανθρωπολόγος Rudolf Virchow (1821-1902) εδραιώνει την κυτταρική θεωρία, αποδεικνύοντας ότι κάθε κύτταρο προέρχεται από ένα άλλο κύτταρο: είναι η γνωστή αρχή “omnis cellula e cellula”. Ο Virchow διενεργεί, το 1871, μεγάλη έρευνα σε 6,7 εκατομμύρια παιδιά στη Γερμανία, για τη σύγκριση των φυσικών χαρακτηριστικών μεταξύ Εβραίων και Χριστιανών. Δεν προκύπτουν διαφορές και η έρευνα δεν έχει κοινωνικό αντίκτυπο. O Virchow είναι ο επιστήμονας που αρθρώνει την τελευταία έγκυρη φωνή στη Γερμανία κατά της άποψης ότι υπάρχουν ανθρώπινες «φυλές» (λίγα μόλις χρόνια αργότερα, το 1883, ο Francis Galton, εξάδελφος του Darwin, επινοεί τον όρο «ευγονισμός», ανοίγοντας τον δρόμο για όλες τις γνωστές αποτρόπαιες συνέπειες της δήθεν εφαρμογής της ευγονικής από τα μετέπειτα ολοκληρωτικά καθεστώτα στη Γερμανία και σε πολλές άλλες χώρες).
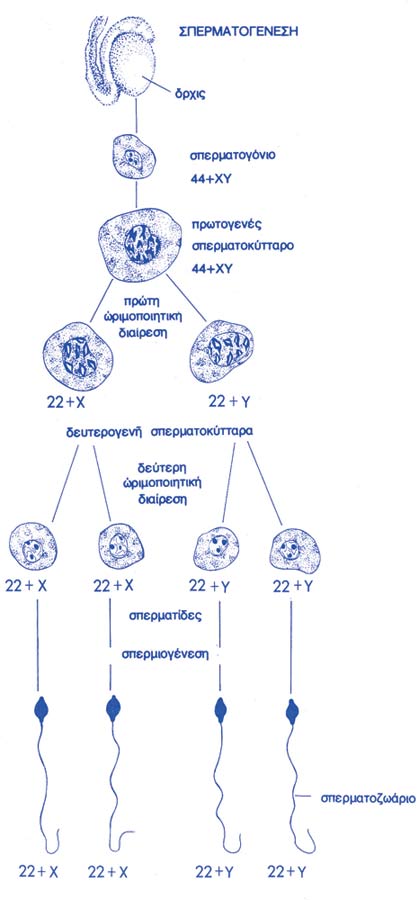
Στα τέλη του 19ου αιώνα, ο Ελβετός ανατόμος και φυσιολόγος Rudolf Albert von Kölliker (1817-1905) ανακαλύπτει τις περίπλοκες διεργασίες που επιτελούνται στον όρχι και αποδεικνύει ότι τα σπερματοζωάρια είναι η κατάληξη μιας μακράς σειράς διαδοχικών κυτταρικών διαιρέσεων (σπερματογένεση), ενώ ο Ελβετός βοτανολόγος Karl Wilhelm von Nägeli (1817-1891) περιγράφει τις φάσεις της μιτώσεως, σε φυτικά κύτταρα, και διαπιστώνει την ύπαρξη του κυτταροπλάσματος, αποτυγχάνει όμως να αντιληφθεί τη σημασία των νόμων του Mendel (βλ. κατωτέρω) και τη σχέση τους με τα χρωμοσώματα. Η ώρα της γενετικής δεν έχει σημάνει ακόμη: ένας σχεδόν άγνωστος Ελβετός χημικός, ο Johannes Friedrich Miescher (1811-1887) ανακαλύπτει τα νουκλεϊκά οξέα (1871), μακρομόρια τα οποία ουδείς θεωρεί άξια λόγου. Λίγα μόλις χρόνια νωρίτερα (1865-6), ο Αυστριακός μοναχός και βοτανολόγος Johann Gregor Mendel (1822-1884) διατυπώνει τους νόμους της κληρονομικότητος (“Versuche uber Pflanzenhybriden”). Η ανακάλυψη περνά απαρατήρητη από τον von Nägeli και τους άλλους ειδικούς της εποχής. Δεκαετίες αργότερα (1900), ο Ολλανδός βοτανολόγος και γενετιστής Hugo de Vries (1848-1935) ανακαλύπτει και ερμηνεύει εκ νέου τους νόμους του Mendel, εισάγει μάλιστα και την έννοια της μεταλλάξεως. Σημειωτέον ότι παράλληλα και ανεξάρτητα, οι νόμοι του Mendel ανακαλύπτονται από τον Γερμανό βοτανολόγο Karl Erich Correns (1864-1933) και τον Αυστριακό αγρονόμο Erich Tschermak von Seysenegg (1872-1962).
Τα τμήματα της σύγχρονης βιολογίας αρχίζουν να συνθέτουν τη σημερινή, γνώριμη πλέον εικόνα: ο Γερμανός βιολόγος και ανατόμος Heinrich Wilhelm von Waldeyer (1836-1921) εισάγει τον όρο «χρωμόσωμα» το 1888 (η πρώτη ανακοίνωση για αυτά τα «σώματα που χρωματίζονται» είναι του F. Schneider, 1873. “Untersuchungen über Plathelminthen.” Oberhessischen Gesellschaft für Natur und Heilkunde 14:69-140). Ο Γερμανός βιολόγος Oskar Hertwig (1849-1922) ανακαλύπτει τους προπυρήνες στα γονιμοποιημένα ωάρια (1875), ενώ ο Ελβετός βιολόγος Herman Fol (1845-1892), ανεξάρτητα από τον Hertwig, παρατηρεί (1879) τη διείσδυση του σπερματοζωαρίου. Ο Βέλγος ζωολόγος Édouard van Beneden (1846-1910) και ο Γερμανός βιολόγος Theodor Boveri (1862-1915) περιγράφουν (1883) την πλοειδία και τη σειρά των σταδίων της μειώσεως. Ο Γερμανός ανατόμος Walther Flemming (1843-1905) χρησιμοποιεί χρωστικές για τη μελέτη των κυττάρων και εισάγει όρους που χρησιμεύουν ακόμη και σήμερα (χρωματίνη, μίτωση, αστέρας…).
Ο Γερμανός φυσιολόγος Johannes Müller (1801-1858), ο οποίος ασχολήθηκε κυρίως με τη νευροφυσιολογία, θεωρείται ως ο πιο σημαντικός φυσιολόγος του πρώτου μισού του 19ου αιώνα. Ένας από τους μαθητές του έμελλε να γράψει ιστορία: ο Γερμανός εξελικτικός βιολόγος και εμβρυολόγος Ernst Heinrich Haeckel (1834-1919, μαθητής των von Kölliker, Müller, Virchow και δάσκαλος των Driesch, Leydig, Hertwig, Roux κ.λπ.), δεχόμενος ισχυρή επιρροή από τον Darwin, αποδέχεται τη θεωρία της εξέλιξης, αλλά διατηρεί επιφυλάξεις ως προς την ορθότητα της φυσικής επιλογής. Η πιο διάσημη άποψή του είναι ότι «η οντογένεση ανακεφαλαιώνει τη φυλογένεση», ενώ εισήγαγε και τον όρο «οικολογία». Ο λεγόμενος αυτός «Νόμος της Βιογενετικής» (1866), εάν ίσχυε, θα σήμαινε ότι το έμβρυο επαναλαμβάνει τα στάδια της εξέλιξης κατά την ανάπτυξή του. Έπεται μεγάλη διαμάχη με τον Virchow, ο οποίος δεν πίστευε στη δαρβινική θεωρία. Ο Haeckel, προκειμένου να εδραιώσει την άποψή του, παραποιεί σκοπίμως σχέδια πρώιμων εμβρύων σε συγκριτικούς πίνακες, αφήνοντας πίσω του το πρώτο γνωστό μελανό παράδειγμα επιστημονικής απάτης. Αργότερα, οι θεωρίες του για τη βάση της εξέλιξης του ανθρώπου σε φυλές χρησιμοποιήθηκαν ως το ψευδο-επιστημονικό υπόβαθρο του Ναζισμού.
Στα τέλη του 19ου αιώνα οι ανακαλύψεις φαίνονται ανεξάντλητες: ο Ιταλός φυσιολόγος και ιστολόγος Enrico Sertoli (1842-1910) ολοκληρώνει την τελική διατύπωση όλων των φάσεων της σπερματογενέσεως, ενώ ο Γερμανός ζωολόγος και συγκριτικός ανατόμος Franz von Leydig (1821-1908) περιγράφει το κύτταρο που φέρει το όνομά του και εκκρίνει τεστοστερόνη. Είναι άξιο περιέργειας ότι η γαλλική σχολή της εποχής, αν και εξαιρετικά ανεπτυγμένη σε άλλους τομείς, δεν εκδηλώνει κανένα σχεδόν ενδιαφέρον για την αναδυόμενη κυτταρική και αναπτυξιακή βιολογία. Για παράδειγμα, ο φυσιολόγος Claude Bernard (1813-1878), πατέρας της Πειραματικής Φυσιολογίας (με το πασίγνωστο σύγγραμμά του “Introduction à l’étude de la médecine expérimentale”, 1865), ουδέποτε ασχολήθηκε με την αναπαραγωγή. Το ίδιο και ο πολυτάλαντος χημικός Louis Pasteur (1822-1895), ο οποίος, ως γνωστόν, άφησε τεράστιο και πολύπλευρο έργο πίσω του σε άλλους τομείς: στην κρυσταλλογραφία, με την οποία έθεσε τις βάσεις της στερεοχημείας, εδραιώνοντας την άποψη ότι η μοριακή ασυμμετρία είναι χαρακτηριστικό της οργανικής ύλης, στη ζύμωση, με την ανακάλυψη της αναερόβιας ζωής, στην τεχνική της παστερίωσης (ο Pasteur απέδειξε, το 1862, ότι η θεωρία της αυτόματης δημιουργίας ήταν λάθος: η αποστείρωση με βρασμό καταργεί την ανάπτυξη των μικροοργανισμών, κάτι που ο Spallanzani είχε ήδη επιτύχει, χωρίς να γίνει πιστευτός στην εποχή του), ή ακόμη στη θεωρία της μικροβιακής προελεύσεως των λοιμωδών νοσημάτων (από το 1865 και τον πρώτο εμβολιασμό ανθρώπου με το αντιλυσσικό εμβόλιο έως τη δημιουργία του Ινστιτούτου Pasteur, το 1886).
Η εμβρυολογία παραμένει εν πολλοίς «γερμανική υπόθεση». Ο Γερμανός εξελικτικός βιολόγος August Friedrich Leopold Weismann (1834-1914) εισάγει τη θεωρία του νεοδαρβινισμού, γνωστή ως «συνθετική» θεωρία, στην οποία διατυπώνεται για πρώτη φορά η σχέση της βιολογίας με τη γενετική (1882-1883): η φυσική επιλογή δρα σε απόκρυφα στοιχεία των γεννητικών κυττάρων, ενώ όλες οι μεταβολές που οφείλονται στο περιβάλλον είναι παροδικές και χάνονται με τον θάνατο του ατόμου. Η κληρονομικότητα οφείλεται σε ουσίες με ειδική δομή, που βρίσκονται στον πυρήνα των γεννητικών κυττάρων, τα οποία είναι δυνητικώς «αθάνατα» και είναι τα μόνα που μεταφέρουν την κληρονομικότητα. Η πειραματική απόδειξη της θεωρίας του Weismann αποτελεί υπόδειγμα, που διδάσκεται ακόμη και σήμερα σε όλα τα συγγράμματα βιολογίας: η αποκοπή της ουράς 901 ποντικών επί 19 διαδοχικές γενεές οδηγεί στην παρατήρηση ότι μέχρι και η τελευταία γενεά διαθέτει ουρές του ιδίου μήκους με την αρχική. Η συνθετική θεωρία έχει ως άμεση επίπτωση την υποχρεωτική διάκριση μεταξύ σωματικών κυττάρων (“soma”) και γεννητικών κυττάρων (“germen”). Απορρίπτονται έτσι οριστικά οι απόψεις του Lamarck περί κληρονομικότητος των επίκτητων χαρακτηριστικών. Σήμερα, βάσει αυτών των απόψεων, διαχωρίζουμε δύο σειρές χαρακτηριστικών για το κάθε είδος: τον «φαινότυπο» (εκδηλούμενη εμφάνιση, ενδεχομένως επίκτητη) και τον «γονότυπο» (γενετικό υλικό που αποτελεί τη βάση της κληρονομικότητος). Οι απόψεις του Weismann οδηγούν στην εκ νέου ανακάλυψη των νόμων του Mendel, τους οποίους ο ίδιος δεν γνώριζε.
O σημαντικός Γερμανός ανατόμος και εμβρυολόγος Wilhelm Roux (1850-1924) εισάγει την «παρεμβατική» ή «πειραματική» εμβρυολογία: η πειραματική παρέμβαση στην εμβρυϊκή ανάπτυξη παρέχει πληροφορίες για τις φυσιολογικές αναπτυξιακές διεργασίες. Ο Roux διεξάγει, το 1888, ένα σημαντικό πείραμα καταστροφής ενός από τα δύο πρώτα βλαστομερίδια εμβρύου βατράχου και παρατηρεί ότι αναπτύσσεται μόνον ένα ημι-έμβρυο. Ο Roux συμπεραίνει, σύμφωνα με την υπόθεση του Weismann, υπάρχει προκαθορισμός των δύο βλαστομεριδίων. Η ερμηνεία είναι λάθος, διότι στην πραγματικότητα υπάρχει επαγωγή από εναπομείναντα στοιχεία του νεκρού κυττάρου, όπως αποδεικνύεται από τον J. F. McClendon το 1910: ο διαχωρισμός των δύο βλαστομεριδίων οδηγεί σε πλήρη ανάπτυξη δύο χωριστών εμβρύων. Έρχεται πλέον η ένδοξη περίοδος της πειραματικής εμβρυολογίας (ή της «αναπτυξιακής μηχανικής» (Entwicklungsmechanik), θεμελιωτές της οποίας θεωρούνται ο Roux και ο Γερμανός βιολόγος και μορφολόγος Hans Driesch (1867-1941). Ο Driesch, με ένα καθοριστικό πείραμα απομόνωσης των βλαστομεριδίων εμβρύου αχινού (1892), αποδεικνύει ότι το εξελικτικό δυναμικό του βλαστομεριδίου είναι μεγαλύτερο από τον εμβρυολογικό προκαθορισμό του, άρα, ο πυρήνας του πρέπει να αλληλεπιδρά με το κυτταρόπλασμα. Σύμφωνα με το πείραμα αυτό, κάθε βλαστομερίδιο αναπτύσσεται σε πλήρη προνύμφη, αν και μικρότερη της φυσιολογικής, γεγονός που αποδεικνύει ότι η ανάπτυξη υπόκειται σε ρύθμιση. Μην μπορώντας να αποδείξει ποιες φυσικές δυνάμεις καθοδηγούν την ανάπτυξη, ο Driesch διακόπτει τον πειραματισμό, γίνεται καθηγητής φιλοσοφίας και ασπάζεται την Αριστοτελική άποψη περί «ενδελέχειας» (ότι δηλαδή κάποια «εσώτερη ζωτική δύναμη» καθοδηγεί την ανάπτυξη των εμβρύων). Αν και δεν ήταν Εβραίος, ο Dreisch υπήρξε ένας από τους πρώτους επιστήμονες που υποχρεώθηκαν σε παραίτηση όταν οι Ναζί ανέλαβαν την εξουσία.
Η πειραματική εμβρυολογία φθάνει στο απόγειό της με το έργο του Γερμανού βιολόγου Hans Spemann (1869-1941) και της βοηθού του Hilde Proescholdt-Mangold (1898-1924). Η μελέτη των μηχανισμών αναπτύξεως του εμβρύου αμφιβίων οδηγεί στην απόδειξη του φαινομένου της επαγωγής και τη διατύπωση της έννοιας του οργανωτή το 1924. Η Hilde Mangold διενεργεί το κρίσιμο πείραμα μεταμόσχευσης ραχιαίου ιστού από ένα έμβρυο σαλαμάνδρας σε άλλο: η μεταμόσχευση προκαλεί την ανάπτυξη δεύτερου, σιαμαίου εμβρύου. (Spemann, Η. Mangold, Η. Über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. Arch. mikr. Anat. und Entw. mech. 100:599-638). Για την εργασία αυτή, ο Spemann βραβεύεται το 1935 με το βραβείο Nobel Ιατρικής & Φυσιολογίας. Όπως έχει δυστυχώς συχνά συμβεί στην ιστορία της επιστήμης, η πρώτη απόδειξη της επαγωγής ανήκει στην Αμερικανίδα βιολόγο Ethel Browne Harvey (1885-1965), η οποία, το 1909, είχε μεταμοσχεύσει ιστό από την περιοχή του υποστόματος της ύδρας στον κορμό μιας άλλης ύδρας και είχε παρατηρήσει αναγέννηση δεύτερου οργανισμού στη θέση μεταμόσχευσης. Η μεθοδολογία της Harvey ήταν ίδια με εκείνη των Spemann και Mangold, όπως και το αποτέλεσμα, αν και η παρατήρηση είχε γίνει σε ασπόνδυλο οργανισμό. Μάλιστα η Harvey, έχοντας επίγνωση της σημασίας του ευρήματος, είχε αποστείλει τη δημοσίευσή της στον Spemann, αλλά η μελέτη αυτή αποσιωπήθηκε πλήρως και δεν βραβεύθηκε από την επιτροπή των βραβείων Nobel (Biol. Bull. 181:72-80).
Αν εξαιρέσει κανείς αυτές τις σκοτεινές πτυχές της ιστορίας και πέρα από τις μεγάλες ανακαλύψεις του, ο Spemann έχει προσφέρει στην εμβρυολογία αμέτρητες τεχνικές και εργαλεία εμβρυϊκής μικροχειρουργικής. Ήδη από το 1896, απομονωμένος σε σανατόριο για αποθεραπεία από φυματίωση, διαβάζει το βιβλίο του August Weismann και ενθουσιάζεται από την εμβρυολογία. Κατά την περίοδο 1901-1903 χρησιμοποιεί τρίχες από τα μαλλιά της νεογέννητης κόρης του για να διαχωρίσει τα δύο πρώτα βλαστομερίδια εμβρύων σαλαμάνδρας. Όταν η περίσφιξη είναι πλήρης, παράγονται δύο πλήρεις δίδυμοι οργανισμοί: καταπίπτει επομένως η θεωρία του Weismann, σύμφωνα με την οποία τα κύτταρα χάνουν τη γενετική πληροφορία σε κάθε μίτωση. Όταν η περίσφιξη είναι μερική, παράγονται δύο σιαμαία έμβρυα: καταφέρεται έτσι το τελικό κτύπημα στη θεωρία του προσχηματισμού. Ωστόσο, αν το επίπεδο της πρώτης μίτωσης (άρα και της περίσφιξης) χωρίζει τον ζυγώτη σε ραχιαία και κοιλιακή πλευρά, μόνον η ραχιαία πλευρά αναπτύσσεται σε πλήρες έμβρυο: άρα, τα κύτταρα καθορίζονται (διαφοροποιούνται) σε κάποια χρονική στιγμή της ανάπτυξης. Την περίοδο 1920-30, ο Spemann μεταφέρει τον πυρήνα ενός βλαστομεριδίου από έμβρυο 16 κυττάρων σε ένα εκπυρηνισμένο βλαστομερίδιο: η κατασκευή εξελίχθηκε σε φυσιολογική σαλαμάνδρα και είχε πραγματοποιηθεί η πρώτη πυρηνική κλωνοποίηση. Το 1936, στο βιβλίο του για την εμβρυϊκή ανάπτυξη και την επαγωγή (“Experimentelle Beiträge zu einer Theorie der Entwicklung”, Springer), ο Spemann προτείνει και το «φανταστικό πείραμα» της κλωνοποίησης ανώτερου οργανισμού από διαφοροποιημένο κύτταρο, ή και ακόμη και από κύτταρο του ενήλικα. Η σχετική τεχνολογία δεν υπήρχε τότε, αλλά αυτό έκτοτε έχει επιτευχθεί από κύτταρο εμβρύου στον βάτραχο (Robert Briggs και Thomas King, 1952) και στα ανώτερα θηλαστικά (Steen Willadsen σε πρόβατο και αγελάδα, το 1984 και 1985 αντίστοιχα). Το πιο πρόσφατο επίτευγμα, με το οποίο ανοίγει και ο δρόμος για πολύ σημαντικές εφαρμογές της πειραματικής εμβρυολογίας στον 21ο αιώνα, είναι η κλωνοποίηση του προβάτου Dolly από κύτταρο ενηλίκου (Ian Wilmut και Keith Campbell, 1995).
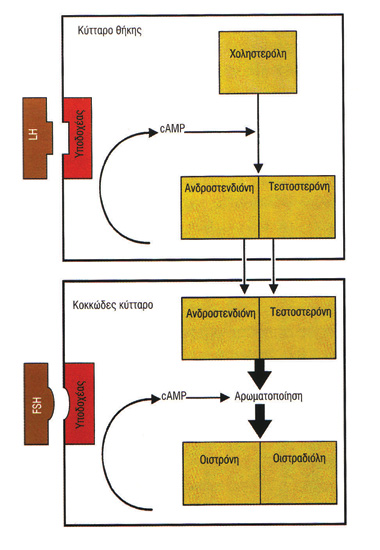
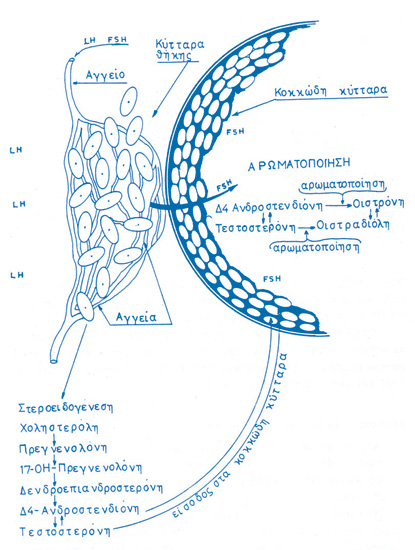
Στις αρχές της δεκαετίας του 1930 ανακαλύπτονται οι ορμόνες του εμμήνου κύκλου και η σχέση τους με την ωοθυλακιορρηξία. Μεγάλο μέρος του έργου αυτού οφείλεται στον Γερμανό βιοχημικό Adolf Frederick Johann Butenandt (1903-1995), ο οποίος περιγράφει τη χημική δομή και τη σύνθεση της οιστρόνης (1929), της ανδροστερόνης (1931) και της τεστοστερόνης (1934), βραβεύεται μάλιστα με το βραβείο Nobel Χημείας το 1939, μαζί με τον Leopold Ruzicka (η ναζιστική κυβέρνηση της Γερμανίας τού απαγορεύει να το δεχθεί και το παραλαμβάνει μεταπολεμικώς, το 1949).
Ο 20ός αιώνας σημαδεύεται όμως και από ραγδαία εξέλιξη του τομέα της γενετικής. Ο Βρετανός βιολόγος και γενετιστής William Bateson (1861-1926) περιγράφει τη γενετική σύζευξη και αποδέχεται πρώτος τους Νόμους του Mendel όταν ανακαλύπτονται εκ νέου. Ο Bateson εισάγει τον όρο «γενετική» το 1909, αλλά και τους όρους «αλληλόμορφο», «ζυγώτης» κ.λπ., ενώ αποδέχεται τη χρωμοσωματική θεωρία μόλις το 1922, μετά από μια επίσκεψη στο εργαστήριο του Αμερικανού βιολόγου και γενετιστή Thomas Hunt Morgan (1866-1945). Μαζί με τον Bateson, ο Morgan θεωρείται θεμελιωτής της σύγχρονης γενετικής. Αν και αρχικώς είναι πολέμιος της φυσικής επιλογής και της χρωμοσωματικής θεωρίας της κληρονομικότητος, μέχρι το 1910, ανακαλύπτει τον γενετικό ανασυνδυασμό (crossing over) μεταξύ δύο ομολόγων χρωμοσωμάτων στη μύγα Drosophila melanogaster, επινοεί τη χαρτογράφηση των χρωμοσωμάτων, συμπεραίνει ότι τα γονίδια ακολουθούν το ένα το άλλο πάνω στα χρωμοσώματα, και αποδεικνύει ότι η ποικιλομορφία οφείλεται σε μεταλλάξεις. Συμπορεύεται για λίγα χρόνια με το κίνημα του ευγονισμού (1910-1920), από το οποίο τελικώς αποχωρεί. Ο Morgan βραβεύεται το 1933 με το βραβείο Nobel Ιατρικής και Φυσιολογίας για την ανακάλυψη του ρόλου των χρωμοσωμάτων στην κληρονομικότητα. Ο Ρωσικής καταγωγής βιολόγος και γενετιστής Theodosius Dobzhansky (1900-1975), ο οποίος μεταναστεύει στις Η.Π.Α. το 1927, γίνεται συνεργάτης του Morgan και αποκτά την αμερικανική ιθαγένεια, στο βιβλίο του (“Genetics and the Origin of Species”, 1937) συνδέει την εξέλιξη με τις γενετικές μεταλλάξεις. Ο Αμερικανός μικροβιολόγος Oswald Theodor Avery (1877-1955) αποδεικνύει, το 1944, ότι το DNA είναι το γενετικό υλικό των κυττάρων: οι μολυσματικές ιδιότητες μεταφέρονται από έναν πνευμονιόκοκκο σε άλλον μέσω της μεταφοράς DNA. Ο Αυστριακός χημικός Erwin Chargaff (1905-2002) μεταναστεύει στη Νέα Υόρκη το 1935 και διατυπώνει τους κανόνες της αναλογίας νουκλεοτιδίων στο DNA το 1950. To 1952 συναντά στο Cambridge τον James Dewey Watson (1928-) και τον Francis Harry Compton Crick (1916-2004) και τους εκθέτει τα ευρήματά του, βάσει των οποίων εκείνοι ανακαλύπτουν τη στερεοχημική δομή του DNA σε διπλή έλικα, το 1953. Η ανακάλυψη αυτή θεωρείται από πολλούς ότι έχει τη σημασία της ανακάλυψης των νόμων της βαρύτητας και δικαίως βραβεύεται με το βραβείο Nobel το 1962. Ωστόσο, δεν βραβεύονται ούτε η Rosalind Franklin, η οποία παρείχε τα κρυσταλλογραφικά δεδομένα που δημοσιεύθηκαν ταυτόχρονα με τη μελέτη των Watson και Crick, ούτε ο Erwin Chargaff, ο οποίος ουσιαστικά τους άνοιξε τον δρόμο. Μόλις μισόν αιώνα αργότερα, στις ημέρες μας, έχει ήδη αναλυθεί σχεδόν πλήρως ολόκληρο το ανθρώπινο DNA (στο πλαίσιο του παγκόσμιου ερευνητικού προγράμματος «ανθρώπινο γονιδίωμα»), αλλά και το γονιδίωμα πολλών άλλων οργανισμών. Αυτό ανοίγει τον δρόμο για συγκριτικές μελέτες, οι οποίες ήδη αρχίζουν να φωτίζουν τους μοριακούς μηχανισμούς της εξέλιξης των ειδών.
Από τη μεταπολεμική αυτή εποχή και μετά, ορισμένοι θεώρησαν ότι η εμβρυολογία έχει ενταχθεί και «ανήκει» πλέον στη γενετική. Η λανθασμένη αυτή άποψη αλλάζει στις μέρες μας, κυρίως με την επάνοδο στο προσκήνιο των μικροχειρισμών, που οδήγησαν στην εξωσωματική γονιμοποίηση και στην κλωνοποίηση. Στην πραγματικότητα, οι σκέψεις αυτές είναι πολύ παλαιότερες της σημερινής γενετικής. Πράγματι, πρώτος ο Schenk (1880) είχε αποπειραθεί να γονιμοποιήσει ωάρια in vitro (στο κουνέλι), χωρίς επιτυχία, ενώ ο Heape (1890) πραγματοποίησε την πρώτη εμβρυομεταφορά στο ίδιο πειραματόζωο. Το 1912 οι Mark και Long προσπαθούν να καλλιεργήσουν ωάρια ποντικού, ενώ η αυλάκωση εμβρύων κουνελιού in vitro επιτυγχάνεται το 1929 (Lewis, Gregory). Η καλλιέργεια εμβρύων επίμυος (Defrise 1933), μακάκου Rhesus (Lewis, Hartman, 1933) μέχρι τα 8 κύτταρα, δημιούργησε ελπίδες για επιτυχή εξωσωματική γονιμοποίηση ωαρίων ποντικού (Lewis, Wright, 1935) και ανθρώπου (Pincus, 1939). Οι προσπάθειες αυτές απέβησαν άκαρπες, μέχρι το 1944, οπότε καταγράφεται (Rock, Menkin) η πρώτη επιτυχής εξωσωματική γονιμοποίηση ανθρώπινου ωαρίου (και διαίρεση σε 2 κύτταρα). Το 1949 ανακοινώνεται η πρώτη παρατήρηση της αυλάκωσης in vitro εμβρύων ποντικού (Hammond) και κουνελιού μετά από εξωσωματική γονιμοποίηση (Chang), ενώ η αυλάκωση in vitro εμβρύων αγελάδος μέχρι τα 24 κύτταρα παρατηρείται το 1951 (Pincus).
Από τη δεκαετία του 1950 μέχρι και σήμερα, η βελτίωση των καλλιεργητικών μέσων είναι διαρκής. Οι πρώτες απόπειρες καθορισμού των αναγκών του εμβρύου in vitro έγιναν το 1956 (Whitten) και μόλις το 1963 ο Brinster επιτυγχάνει την καλλιέργεια εμβρύων ποντικού (μέχρι του σταδίου της βλαστοκύστης), σε χημικώς καθορισμένο μέσον. Στη συνέχεια, πάμπολλοι σύγχρονοί μας ερευνητές ασχολούνται με το θέμα αυτό (Biggers, Brinster, Bavister, Chang, Daniels, Edwards, Hafez, Wales, Whitten, Whittingham, Yanagimachi) και αναπτύσσουν μέσα για την καλλιέργεια εμβρύων πολλών θηλαστικών. Ένας εξ αυτών, ο Robert G. Edwards, έμελλε να γίνει ο πιο διάσημος βιολόγος της εποχής του, όταν ανακοίνωνε, στις 25 Ιουλίου του 1978, τη γέννηση της Louise Brown, του πρώτου «παιδιού του σωλήνα», στην Αγγλία, από κοινού με τον χειρουργό μαιευτήρα Patrick Steptoe και τη μαία Jane Purdie (Εικ. 26).
Το επίτευγμα αυτό θα ήταν αδύνατο χωρίς την ανάπτυξη της λαπαροσκοπικής χειρουργικής (Kelling, Jacobeus που εισήγαγε τη γυναικολογική θέση trendelenburg), απόρροια η ίδια διαφόρων άλλων τεχνολογικών επιτευγμάτων που πέρασαν σχεδόν απαρατήρητα, όπως είναι η επινόηση της ψυχρής φωτεινής πηγής (Hopkins), ή η θεαματική βελτίωση των αναισθητικών μεθόδων και των χειρουργικών εργαλείων. Οι παλαιότεροι ενθυμούνται ότι η εξωσωματική γονιμοποίηση, στις πρώτες ημέρες της, προϋπέθετε μια λαπαροσκοπική επέμβαση για τη συλλογή των ωαρίων, ότι δεν υπήρχαν φάρμακα για τη διέγερση της ωοθηκικής λειτουργίας, ενώ δεν υπήρχε τρόπος να προγραμματισθεί η ωοθυλακιορρηξία, με συνέπεια ολόκληρη η ιατροβιολογική ομάδα να πρέπει να αναμένει επί ώρες την κατάλληλη στιγμή της ωοληψίας. Σημειωτέον ότι η ίδια η υπερηχοτομογραφία εισήχθη, ως τεχνική, μόλις το 1958 (Donald), η δε διακολπική παραλλαγή της μια δεκαετία αργότερα (Kratochwil, 1967), ενώ η εφαρμογή της διακολπικής τεχνικής στην ωοληψία χρονολογείται μόλις στα 1983 (Wikland, Fertil Steril 39:603).
Έκτοτε, οι εξελίξεις υπήρξαν ραγδαίες: στον τομέα της κρυοβιολογίας, η πρώτη γέννηση από κρυοσυντηρημένο έμβρυο επιτυγχάνεται το 1983 (Trounson), το 1984 (Renard, Testart) εισάγεται η χρήση της προπανδιόλης ως κρυοπροστατευτικού μέσου, το 1985 (Cohen) ανακοινώνεται η πρώτη επιτυχής κατάψυξη ανθρώπινης βλαστοκύστεως και το 1986 (Lassalle) η πρώτη επιτυχής κατάψυξη ανθρώπινου ζυγώτη (στάδιο προπυρήνων). Στον τομέα της αντιμετώπισης του ανδρικού παράγοντα υπογονιμότητος, προτείνονται μέθοδοι μικροκαλλιέργειας (Ord, 1990), και σημειώνεται επανάσταση όταν ανακοινώνεται η επιτυχής μικρογονιμοποίηση με ενδοωαριακή έγχυση σπερματοζωαρίου (ICSI) το 1992 (Palermo) και οι παραλλαγές της, MESA (Silber & Asch, 1992) και TESE (Schoyssman, 1993). Ο ανδρικός παράγων υπογονιμότητος θεωρείται ότι έχει πρακτικά εξαλειφθεί, μετά την ανακοίνωση της χρήσης σπερματίδων στη μικρογονιμοποίηση (Fishel, Tesarik, 1995). Παράλληλα, αναπτύσσονται νέες τεχνολογίες, εκ των οποίων οι πλέον σημαντικές θεωρούνται η απομόνωση σειρών βλαστικών εμβρυϊκών κυττάρων (Evans & Kaufman, 1981), η κατασκευή διαγονιδιακών ζώων (Gordon, Brinster, Palmiter, 1983), η προεμφυτευτική γενετική διάγνωση (Handyside, 1989), καθώς και η υποβοηθούμενη εκκόλαψη και η μετάγγιση κυτταροπλάσματος ωαρίου (Cohen, 1999). Ένα τελευταίο σημαντικό επίτευγμα, το οποίο, σε συνδυασμό με την αυξημένη βιολογική τεχνογνωσία, έχει επιτρέψει τη σταθεροποίηση των ποσοστών επιτυχίας της εξωσωματικής γονιμοποίησης σε υψηλά επίπεδα την τελευταία οκταετία, είναι η κατασκευή συνθετικών γοναδοτροπινών, μέσω της τεχνολογίας του ανασυνδυασμένου DNA.
Σήμερα πια, ο γυναικολόγος και ο εμβρυολόγος διαθέτουν μια πανοπλία μεθόδων για την κλινική αντιμετώπιση της υπογονιμότητος (ιατρικώς υποβοηθούμενη αναπαραγωγή), ενώ παράλληλα η βασική έρευνα στην εμβρυολογία αναμένεται να φέρει και νέα επανάσταση: τη γενίκευση της χρήσεως των εμβρυϊκών βλαστικών κυττάρων στην ιατρική του 21ου αιώνα για τη θεραπεία δεκάδων παθήσεων, μέχρι σήμερα ανίατων, αλλά και για την αναδυόμενη αναγεννητική ιατρική (δηλαδή τη δημιουργία ιστών και οργάνων από βλαστικά κύτταρα για αυτόλογη χρήση). Η εμβρυολογία, σε συνδυασμό με τη σύγχρονη γενετική και την αναπτυξιακή βιολογία, βρίσκεται σε μια εξαιρετικά ενδιαφέρουσα καμπή της ιστορίας της, αφού σύντομα αναμένεται να εξελιχθεί στο κεντρικό επιστημονικό υπόβαθρο της βιοϊατρικής του μέλλοντος.








.")
.")
.")
.")
")


 ωοθυλάκιο.")




















.")
.")